Азот в растениеводстве
Азот в растениеводстве
Азот — химический элемент, инертный газ без цвета и запаха, открыт французским химиком Лавуазье во второй половине XVIII в., является основным компонентом атмосферного воздуха (78,08%). Название означает «нежизненный», так как не поддерживающий горение и дыхание. Однако, дальнейшие исследования показали огромную роль азота в жизни растений и всего органического мира.
Азот входит в состав:
- белков, пептидов и аминокислот, которые являются составной частью протоплазмы и ядра растительных клеток;
- нуклеиновых кислот (ДНК и РНК) — носителей наследственных свойств живых организмов и участвующих в обмене веществ;
- молекул хлорофилла;
- ферментов;
- фосфатидов;
- гормонов;
- большинство витаминов.
Азотное питание растений
Все ферменты — белковые вещества, поэтому при недостаточном снабжении растений азотом синтез ферментов замедляется, что приводит к нарушениям в процессах биосинтеза, обмена веществ, в итоге, к снижению урожая.
Регулирование азотного питания растений, можно влиять на урожайность сельскохозяйственных культур с учетом других факторов жизни. Максимальный урожай достигается при достаточном обеспечении растений всеми условиями их роста. Академик Д.Н. Прянишников писал, что вся история земледелия в Западной Европе говорит о том, что главным условием, определяющим среднюю высоту урожаев в разные эпохи, была степень обеспеченности сельскохозяйственных растений азотом.
Оптимальное азотное питание способствует синтезу белковых веществ, растения образуют мощные стебли и листья с интенсивной зеленой окраской. Мощный ассимиляционный аппарат позволяет накапливать большее количество продуктов фотосинтеза, повышая урожайность и, как правило, его качество.
Одностороннее избыточное питание азотом, особенно во второй половине вегетационного периода, приводит к задержке созревания растений; образуется большая вегетативная масса, урожай репродуктивных органов но не успевает сформироваться.
Недостаток азота приводит к сильному замедлению роста растений. Прежде всего сказывается на развитии вегетативной массы: листья становятся мелкими, светло-зелеными, раньше желтеют, стебли тонкие, слабо ветвятся. Снижается формирование репродуктивных органов, урожай резко снижается. Азотное голодание у злаковых культур приводит к ослаблению кущения, уменьшается количество зерен в колосе, снижается белковость зерна.
Содержание азота в растениях
По химическому составу, на долю азота в растениях приходится 0,5-5,0% воздушно-сухой массы, основное количество приходится на семенах. Содержание белка четко коррелирует с количеством азота в растениях. В вегетативных органах содержание азота ниже: в соломе бобовых 1,0-1,4%, в соломе злаковых 0,45-0,65%. Еще меньше азота накапливается в корне-, клубнеплодах и овощных культурах: картофель (клубни) 0,32%, сахарная свекла (корни) 0,24%, капуста 0,33% сырого вещества.
Таблица. Содержание белка и азота в семенах различных культур, %
| КУЛЬТУРА | БЕЛОК | АЗОТ |
|
Соя |
29 | 5,8 |
|
Горох |
20 | 4,5 |
|
Пшеница |
14 | 2,5 |
|
Рис |
7 | 1,2 |
Содержание азота в растениях зависит от возраста, почвенно-климатических условий, питательного режима, в частности обеспеченности питательными элементами.
Содержание азота в молодых вегетативных органах выше. По мере старения азотистые вещества мигрируют в появляющиеся листья и побеги.
Таблица. Содержание азота в вегетативной массе зерновых культур по фазам развития, % на воздушно-сухое вещество
| КУЛЬТУРА | ФАЗА РАЗВИТИЯ | |||
| кущение | трубкование | колошение | цветение | |
| Озимая пшеница | 5,0-5,4 | 3,0-4,5 | 2,1-2,5 | 2,0-2,4 |
| Яровая пшеница | 4,5-5,5 | 3,0-4,4 | 2,5-3,0 | 1,8-2,5 |
| Овес | 5,5-5,9 | 2,9-3,9 | 2,2 | 1,3-1,7 |
Поступление и трансформация азота в белковые вещества
Темпы накопления органических веществ растениями опережают поступление азота и других питательных веществ. Происходит «ростовое разбавление» содержания питательных элементов. При созревании отмечается выраженное передвижение азота в репродуктивные органы, где они накапливаются в виде запасных белков.
В основном азот поступает в растения в нитратной и аммонийно форме, но также способны усваивать некоторые растворимые органические соединения, например, мочевину, аминокислоты, аспарагин.
Из поступающих из почвы в растения соединений азота только аммиак непосредственно используется для синтеза аминокислот. Нитраты и нитриты включаются в синтез аминокислот только после восстановления в тканях растений.
Редукция нитратов до аммиака начинается уже в корнях с помощью флавиновых металлоферментов:
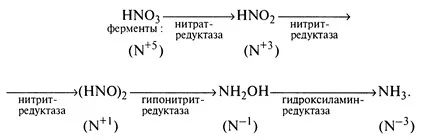
При избытке, часть нитратов поступает в неизменном виде в листья, где восстанавливается по той же схеме.
Образование аминокислот (аминирование) происходит в результате взаимодействия аммиака с кетокислотами: пировиноградной, щавелевоуксусной, кетоглутаровой и др., образующиеся в процессе окисления углеводов. Аминирование регулируется ферментами. Так, при взаимодействии пировиноградной кислоты с аммиаком образуется аланин:
![]()
Аналогично взаимодействие аммиака с щавелевоуксусной кислотой приводит к образованию аспарагиновой кислоты (СООН-СН2-СНNН2-СООН), с кетоглутаровой кислотой — глутаминовая кислота (СООН-СН2-СН2-СНNН2-СООН).
В аминокислоты азот входит в виде аминогруппы (- NH2). Процессы образования аминокислот происходит в корнях и в надземной части растений.
Опыты с использованием меченых атомов показывают, что уже через несколько минут после подкормки растений аммиачными удобрениями, в тканях могут обнаруживаться аминокислоты, синтезированные из внесенного в подкормку аммиака. При этом первой образующейся аминокислотой является аланин, затем аспарагиновая и глутаминовая кислоты.
Нитратный азот может накапливаться в растениях в больших количествах, без причинения им вреда. Аммиак в свободном виде в тканях содержится в незначительных количествах. Его накопление, особенно при недостатке углеводов, приводит к аммиачному отравлению растений.
Однако растения имеют способность связывать избыток свободного аммиака: его часть вступает во взаимодействие с синтезированными аспарагиновой и глутаминовой аминокислотами, образуя соответствующие амиды — аспарагин и глутамин:
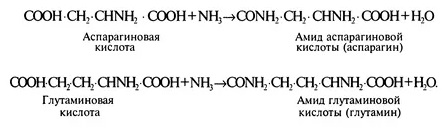
Образование аспарагина и глутамина позволяет растениям защитить себя от аммиачного отравления и создать резерв аммиака, кроме того, амиды участвуют в синтезе белков.
В 1937 г. биохимиками А.Е. Браунштейном и М.Г. Крицманом была открыта реакция переаминирования, заключающаяся в переносе аминогруппы с аминокислоты на кетокислоту с образованием других амино- и кетокислот. Реакция катализируется ферментами трансаминазами или аминоферазами.
Так, присоединение к пировиноградной кислоте аминной группы от глутаминовой кислоты, приводит к образованию аланина и кетоглутаровой кислоты:
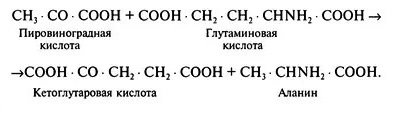
Благодаря переаминированию синтезируется значительное число аминокислот. В растениях наиболее легко переаминируются глутаминовая и аспарагиновая кислоты.
Аминокислоты являются составными частями полипептидов и белков. В построении белковых молекул участвуют 20 аминокислот, аспарагин и глутамин в различных соотношениях и пространственной ориентации, что обуславливает огромное разнообразие белков. В настоящее время известно более 90 аминокислот, около 70 из них присутствуют в растениях в свободном виде и не входят в состав белков.
Растения синтезируют аминокислоты, которые не могут образовываться в организме человека и высших животных, но являются незаменимыми для их жизни. К ним относятся: лизин, гистидин, фенилаланин, триптофан, валин, лейцин, изолейцин, треонин и метионин.
На долю небелкового органического азота в растениях приходится 20-26% от общего количества. В неблагоприятных условиях, например, при дефиците калия или недостаточном освещении, количество небелковых азотистых соединений возрастает.
В тканях растений белки находятся в динамичном равновесии с небелковыми азотистыми соединениями. Одновременно с синтезом белков и аминокислот протекает процесс их распада: отщепление аминогруппы от аминокислоты с образованием кетокислот и аммиака. Этот процес называется дезаминированием. Высвобождающаяся кетокислота используется растениями для синтеза углеводов, жиров и иных веществ; аммиак повторно вступает в реакцию аминирования других кетокислот, образуя новые аминокислоты, при его избытке — аспарагин и глутамин.
Таким образом, весь цикл превращений азотистых соединений в растениях начинается (аминирование) и заканчивается (дезаминирование) аммиаком.
За все время вегетации растения синтезируется большое количество белковых соединений, причем в разные периоды роста обмен азотистых веществ происходит по-разному.
При прорастании семян, клубней, луковиц наблюдается распад запасных белков. Продукты распада расходуются на синтез аминокислот, амидов и белков в тканях проростков до выхода их на поверхность почвы. В Затем, по мере формирования корневой системы и листового аппарата, синтез белков протекает за счет минерального азота, поглощаемого из почвы.
В молодых растениях преобладает синтез белков. В процессе старения растений начинает преобладать распад белков. Продукты распада из стареющих органов мигрируют в молодые, интенсивно растущие органы, где используются для синтеза новых белков в точках роста. По мере созревания растений и формирования репродуктивных органов, белковых веществ распадаются в вегетативных частей, продукты распада перемещаются в репродуктивные органы, где используются для образования запасных белков. К этому моменту поступление азота в растения из почвы существенно замедляется или полностью прекращается.
Особенности аммонийного и нитратного питания растений
В конце XIX в. в агрономической науке ведущую роль занимала теория нитратного питания растений, роль аммиака как источника минерального питания отрицалась.
Причинами этому послужили:
- опыты в водных культурах: отмечалось хорошее развитие растений на фоне нитратных солей, на фоне аммонийных солей развитие было плохим;
- открытие процесса нитрификации в почве; что стало основанием считать: при внесении в почву аммонийных удобрений они переходят в нитратную форму, которая усваивается растениями;
- внесение чилийской селитры (NaNO3) заметно повышало урожайность культур.
Однако в конце века П.С. Коссович в опытах со стерильными культурами показал, что растения могут также усваивать аммиачный азот без окисления в нитратную форму. К такому же выводы пришел и французский исследователь Мазе в 1900 г. После этого были изучены условия и особенности питания аммонийными и нитратными формами азота. Фундаментальные исследования по этому вопросу провел Д.Н. Прянишников. Он показал, что эффективность использования различных форм азота зависит от реакция среды: в нейтральной реакции лучше поглощается аммонийный азот, при кислой — нитратный.
В начальные фазы роста существенное значение имеют биологические особенности. При прорастании семян с небольшим запасом углеводов, например, у сахарной свеклы, а, следовательно, органических кетокислот, избыточное поступление аммония в растения оказывает негативное действие. Аммонийный азот не успевает использоваться для синтеза аминокислот, накапливается в тканях растения и вызывает их отравление. В данном случае используют нитратные формы азотных удобрений, так как они также накапливаться в тканях растений, но не причиняют вреда. Семена и посевной материал с большим запасом углеводов, например, картофель, используют аммонийный азот для синтеза аминокислот без ограничений. Поэтому для таких культур аммонийная и нитратная формы в начальные стадии роста равноценны.
На поглощение нитратного и аммонийного азота влияет обеспеченность другими элементами питания. Повышенное содержание в почве калия, кальция и магния способствует поглощению аммония. При нитратном питании значение имеет обеспеченность растений фосфором и молибденом. Дефицит молибдена приводит к задержке восстановления нитратов до аммиака и способствует накоплению нитратов в тканях растений.
Учитывая, что аммонийная форма азота при поступлении в растения может сразу использоваться для синтеза аминокислот, тогда как нитратная только после восстановления до аммиака, аммоний более энергетически экономной формой.
Круговорот и баланс азота в земледелии
Минеральные соединения азота в почве подвижны. Их запас и трансформация являются результатом многочисленных, часто взаимозависимых физических, физико-химических и биологических процессов круговорота азота.
В целинных почвах естественных биоценозов протекает замкнутый цикл круговорота азота и других биогенных элементов.
Он включает приходные статьи:
- поступление азота с растительным опадом и остатками корней;
- экскрементами и останками животных;
- биологическую фиксацию атмосферного азота микроорганизмами;
- поступление с атмосферными осадками (образование NO3— и NН4+ под действием грозовых разрядов и промышленные выбросы).
Расходные статьи:
- потребление азота растениями;
- инфильтрация;
- денитрификция;
- потери от водной и ветровой эрозии;
- иммобилизация азота — превращения минерального азота в органический, (в этом случае азот не теряется, а переходит в недоступную для растений форму).
Соотношение приходных и расходных частей в круговороте азота составляет баланс этого элемента. В случае превышения расхода над накоплением он отрицательный, при превышении поступления над расходом — положительный, при равенстве частей баланса — уравновешенный, или нулевой.
Для природных биоценозов обычно характерен уравновешенный баланс азота и других биогенных элементов. Потери от вымывания и денитрификации компенсируются его поступлением с осадками и биологической азотфиксацией.
В результате распашки почв азотный режим претерпевает изменения: расходные части баланса резко возрастают. Большое количество азота отчуждается с сельскохозяйственной продукцией. Интенсивная обработка почвы приводит к усилению минерализации органического вещества, увеличению потерь за счет инфильтрации, денитрификации и эрозии.
В условиях достаточного и избыточного увлажнения, орошения, особенно на легких почвах, увеличивается расход нитратов на вымывание вместе из корнеобитаемого слоя в дренажные воды. Суглинистые и глинистые почвы, богатые гумусом, лучше удерживают воду, поэтому вымывание нитратов незначительное — до 3-5 кг/га нитратного азота в год. На легких по гранулометрическому составу, особенно в паровом поле, потери нитратного азота от вымывания достигают 20-30 кг/га в год.
Потери газообразных форм азота от денитрификации составляют также имеют существенное значение. Процесс денитрификации протекает в анаэробных условиях. Логично предположить, что распашка устраняет эти условия, однако, разрыхленная почва состоит из отдельных структурных агрегатов (комков) разного размера. Внутри каждого почвенного агрегата создаются анаэробные условия, и тем интенсивнее, чем больше непоглощенных растениями нитратов накапливается.





