На каждой свиноферме должно быть 6-8 отдельных больничных загонов на сотню свиноматок. Должно быть доступно по крайней мере два загона для отъемышей, два для производителей, два для откормочных поросят и два загона для хромых или неблагополучных свиноматок.
Приобретенная свинья является наиболее важным потенциальным источником новых инфекций, и, хотя за стадом-донором можно тщательно следить для определения состояния его здоровья, на момент покупки инфекционное заболевание могло находиться в стадии инкубации.
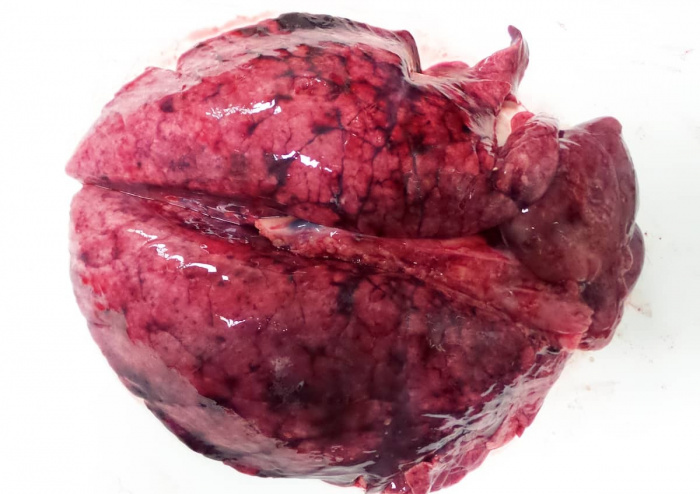
Бактерии, как и вирусы, могут поражать определенные части анатомии или отдельную систему. Например, Actinobacillus pleuropneumoniae поражает плевру (гладкую поверхность, покрывающую легкие) и легочную ткань под ней. E. coli или сальмонелла проникают в тонкий кишечник, вызывая энтерит, но Salmonella choleraesuis, адаптированный к хозяину серотип сальмонелл свиней, может поражать весь организм, включая легкие, вызывая пневмонию.
Основная роль животновода на ферме заключается в оценке взаимодействия свиньи, ее возраста и /или продуктивного цикла с качеством, содержанием и потреблением корма. Роль менеджмента в этом отношении оказывает важное влияние не только на уровень заболеваемости в стаде, но и на то, максимально ли свинья использует свой биологический потенциал.
Решать задачи сбора, учета и анализа данных для организации последующего эффективного кормления помогает специализированное оборудование для откорма свиней.
Оценка состояния тела свиноматки. Измеритель состояния тела свиноматки - это новая экономически эффективная технология, разработанная доктором Кнауэром и его коллегами, которая позволяет объективно оценивать состояние тела.
Диарея после отъема поросят приводит к замедлению роста, снижению производственных показателей и, в худшем случае, к смерти. Кишечная микробиота способствует пищеварению, поддерживает нормальные функции слизистой оболочки кишечника и защищает от патогенов. Во время отъема потеря кишечной микробиоты в слое слизи, защищающем эпителий, может привести к диарее.
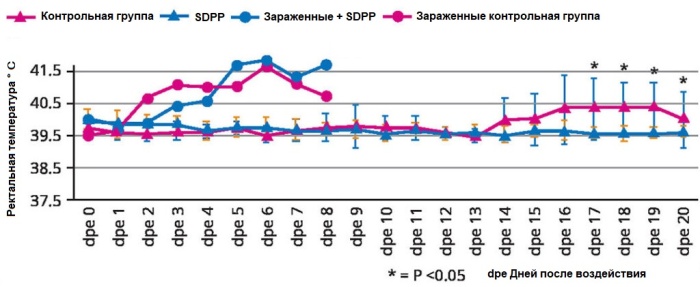
Инструментарий для борьбы с африканской чумой свиней все еще относительно пуст. Наиболее широко используемым подходом являются строгие меры биозащиты, при этом новаторская вакцинация разрешена только в некоторых регионах Азии. Было замечено, что подход к питанию с использованием плазмы позволяет смягчить тяжесть вируса.
Изменения в поведении свиней являются индикаторами проблем со здоровьем и благополучием. Измерение активности свиней может использоваться для обнаружения изменений в поведении. Видеоанализ или пассивные инфракрасные детекторы движения - вот некоторые из методов, используемых для измерения активности свиней в загонах.
Чтобы эти свиньи продолжали работать на уровне своего генетического потенциала, воздействие патогенов на них должно быть сведено к минимуму. Минимальное воздействие патогенов является целью комплексной программы биозащиты.
Исследователи из Нидерландов недавно оценили сочетание сроков группирования и раздельного отлучения от груди для улучшения набора веса поросятами с низким весом при рождении в системах многососного вскармливания.
Исследователи из Тайваня исследовали влияние
модифицированного двухступенчатого ферментированного продукта из пера и соевой муки (TSFP) на продуктивность, клиническую биохимию кр...
Надежное сравнение благосостояния различных
типов свиноводства
Ученые Кембриджского университета разработали
систему измерения благосостояния животных, которая позволяет про...
Посторонний поросенок может вмешаться,
чтобы уменьшить атаки или беспокойство жертвы
Небольшое исследование показывает, что
когда дерутся две свиньи, сторонняя свинья может ...